Ideas, Notes, Curiosities
concerning ecology and evolution of mosquitoes


Figure 1. “The Discovery of Honey by Bacchus.” Piero di Cosimo, ca 1499. Tempera with oil glazes on panel. Worcester Museum of Art. [could this actually be the first sampling of a treehole? Note that some of these folk are satyrs, which makes this illustration relevant to Section III.
Introduction
Communities are so complex that it is exceedingly difficult to study them even when relatively little is known. The situation is exacerbated when a great deal is known. Pianka (2010)
I am about done. If I had another 10-20 years to kill in the lab, here are some of the things I would want to work on, or with which to torture my students. Most of these ideas stem from the work of my students, either published, unpublished, or abandoned in frustration. I offer these thoughts to colleagues in the hope that some may stimulate successful projects. I’d welcome any questions or suggestions and would gladly obtain local (central Massachusetts) samples of mosquitoes for further work. I am posting this dull diatribe here in hopes that it may result in some useful or amusing (if dull) dialogue. Topics include the vertical stratification of habitat choice by two sympatric treehole mosquitoes (Ochlerotatus hendersoni and Oc. triseriatus), miscellaneous thoughts about various other treehole North American treehole species, and the invasion of the Americas by the tiger mosquito, Aedes albopictus. Other topics may be added as time and interest permit. For additional details about things that have happened at Clark University’s Lasry Room 210, please visit our lab web site.
Disclaimer: some of these ideas and speculations may be either naïve or out of date, as I lack any formal entomology training, have never had much patience with the mosquito literature, and have been relatively isolated from good entomology literature sources throughout my career (admittedly poor excuses), especially since retiring in 2017.
I. Stratification.
The Ochlerotatus (formerly Aedes) hendersoni/triseriatus stratification (or lack thereof), in which hendersoni occupies high larval habitats and triseriatus prefers low treeholes. This phenomenon exists in much of the range of the two species (Fig. 2), but not so much in some areas in the southeast; in allopatric parts of hendersoni’s range, the species prefers basal habitats, suggesting that hendersoni has been forced by interactions with triseriatus to prefer higher habitats in areas where the species co-occur (Fig. 3, 4). See the work of my PhD student Jamie Fitzgerald for further background (Fitzgerald and Livdahl 2021). See also the excellent work of Rob Copeland (Copeland and Craig 1990, 1992), which showed that larvae of hendersoni are inferior competitors to those of triseriatus.
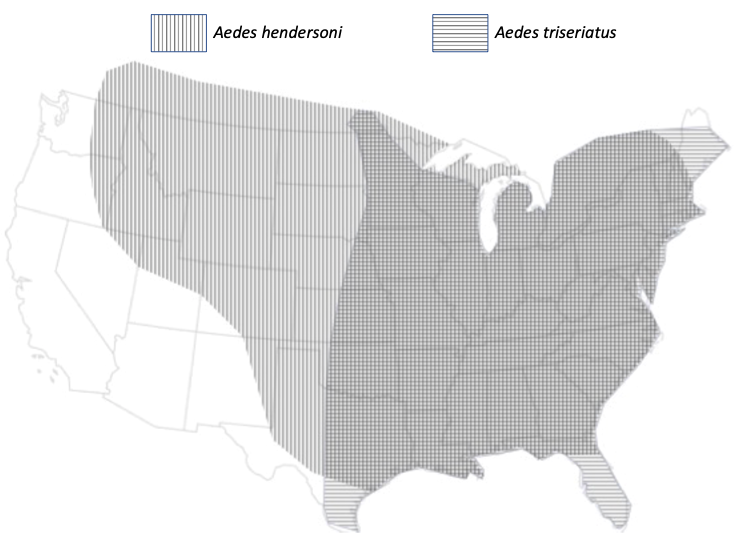
Figure 2. Range maps for Ochlerotatus (=Aedes) triseriatus (horizontal lines) and Oc. hendersoni (vertical lines). From Fitzgerald and Livdahl (2021), modified from Darsie and Ward (2004).
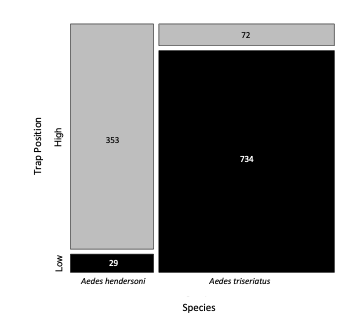
Figure 3. Mosaic plot of trap data for Ochlrtotatus (=Aedes) triseriatus and Oc. hendersoni in sympatric sites, pooled across five locations. Numbers are based on estimated oviposition events. Width of each vertical column indicates relative abundance of each species, and height of each rectangle indicates relative frequency of occurrence within high and low traps.
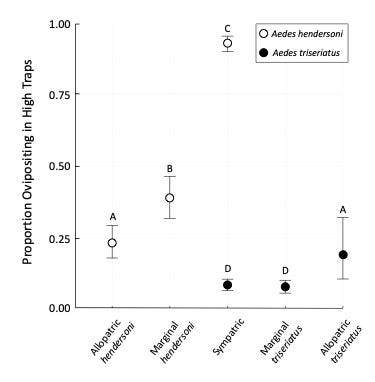
Figure 4. Proportions of oviposition events occurring in high ovitraps, for all species occurrence category and species combinations. The overall proportion, pooled across sites is shown for each group, and 95% confidence intervals are shown for each proportion. Groups that are not significantly different (as indicated by Marascuilo pairwise contrasts) are indicated by common letters below each column. From Fitzgerald and Livdahl (2021).
This stratification phenomenon is reminiscent of the classical studies of intertidal barnacle species competition and stratification by Connell (1960). In the barnacle case, the preferred habitat is the lower intertidal and subtidal, occupied by the superior competitor. Physical stress (high temperature, more desiccation) in the lower intertidal and subtidal is less severe. In the mosquito case, the preferred habitat is basal treeholes, which receive more nutrient input due to more stem flow and are subject to lower desiccation than higher treeholes (this assertion awaits empirical study but seems highly likely). In both cases, the inferior competitor is confined to the low rent district (higher habitats).
Stratification between these mosquitoes is potentially a compelling example of competitive displacement, niche partitioning, resource partitioning, etc. As such, it is of great potential interest to ecologists and evolutionary biologists beyond the narrow confines of mosquito ecology and evolution but remains one of mosquito biology’s best kept secrets. The mechanism for the two species’ preferences and the ultimate factor(s) determining the preferences are not known; further work with this system will be difficult, as the species are not easily distinguished from one another and hendersoni does not mate readily in laboratory cages. Nonetheless, a few questions are worth pursuing:
A. Why are the two species not stratified throughout their sympatric regions? I hypothesize that larvae of filter feeding mosquitoes in regions that are dominated by the predatory Toxorhynchites rutilus do not face intense interspecific competition, as argued by Bradshaw and Holzapfel (1983) in their year-long study of Florida treehole habitats. If not, hendersoni preferences for high habitats may not have been selectively favored.
B. Are there alternatives to competition as factors driving stratification, such as tolerance/virulence of the two Ochlerotatus species to one another’s parasites? We have found that a gregarine parasite can regulate a natural populaton of Aedes albopictus in Bermuda, even though it is not particularly virulent (Soghigian and Livdahl 2021). Ascogregarina taiwanensis, the gregarine of Ae. albopictus, can complete its life cycle within Aedes japonicus Erthal et al. 2012) Gregarines infect both Oc. hendersoni and Oc. triseriatus, and the degree of host specificity, cross-infection and virulence of these possible host-parasite associations is a ripe topic for further work.
C. What is the mechanism for stratification where it occurs, and is it genetically based? Comparative whole genome sequences of both species from sympatric and allopatric regions may reveal chromosomal regions with genes that could account for the habitat preferences. I suggest searching for locations of genes known to influence wing morphology, beat frequency and consequent upward and downward flight in Drosophila (see, e.g., Ray et al. 2016) as a starting point.
Alternative mechanisms may lie in different attractions of the species to olfactory cues, humidity differences in high and low habitats, or light regimes in canopy and basal habitats. These alternatives could be explored experimentally in lab settings.
D. What are the factors that restrict cross-mating between these species? Hybridization does occur, at least here in Massachusetts, but it is not common. Does it occur more commonly in regions where stratification is weaker or non-existent? I would expect that to be the case. It may be worthwhile to examine wingbeat frequencies of the females of these species, as male detection of females could be their initial cue. See Section III.C.2.b.
E. Is the hendersoni/triseriatus species pair an example of sympatric speciation? Perhaps methods of phylogenomics using samples from various parts of the ranges of these species, along with other members of the clade, such as brelandi could shed light on this question, informed by the paleoecology of forests in North America. This is beyond my pay grade, but somebody might have fun with it.
F. Jamie Fitzgerald observed another curiosity, that both these species avoid basal habitats next to a flood-prone river (avoiding the risks of eggs hatching from a submerged treehole into a river during a flood), and that triseriatus predominated in ovitraps high in the trees in such situations (Fig. 5, Fitzgerald and Livdahl 2023). This occurred in two successive years at two distinct locations, despite that the two species stratified with hendersoni laying eggs in high traps and triseriatus in basal traps in nearby upland forests. Is this a local phenomenon or can it be found more generally? Additional follow-up studies in replicated floodplains are necessary. If it is a general phenomenon, further studies would be promising into the mechanisms of 1) avoidance of basal streamside habitats by both species, and 2) the apparent avoidance of higher habitats by hendersoni near streams. I hypothesize a behavioral switch by hendersoni leading to avoidance of competition with triseriatus as perhaps the simplest underlying explanation.
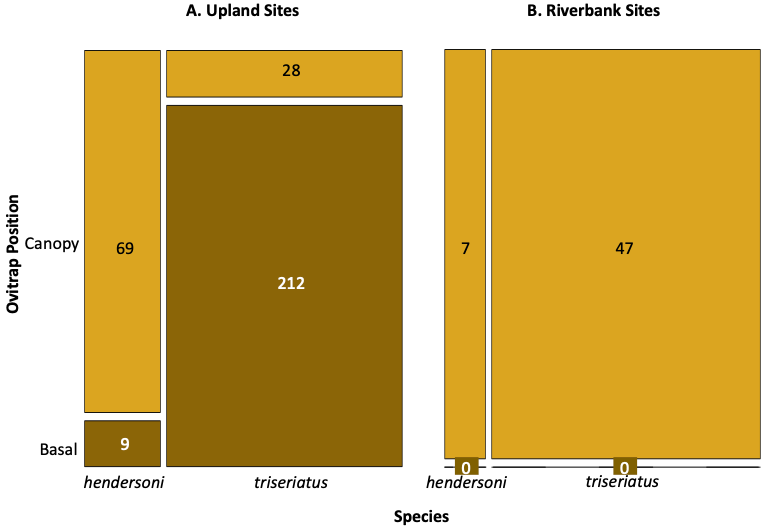
Figure. 5. A: Mosaic plot of trap data for Aedes triseriatus and Ae. hendersoni in upland forested sites at basal and elevated positions. B: trap data from riverbank sites. From Fitzgerald and Livdahl (2023).
II. Questions about other treehole species

Figure 6. Treehole mosquitoes found in central Massachusetts. Painting by Edward Rooks.
A. Anopheles barberi.
1. Anopheles barberi habitat preferences. This might be the most poorly studied anopheline in North America. In my (generally sloppy) sampling efforts, I’ve found it to be sporatic and generally rare, but when I have found it, it’s usually been in rot-holes, i.e., cavities in trees that have a vertical opening. I am reminded of Phil Lounibos’ study of maternal care by African Trichosopon digitatum in which the females guard their eggs (which float on the surface, as do those of A. barberi) against being washed out of cacao husks during heavy rains (Lounibos et al. 1983). I hypothesize that A. barberi’s habitat choice is a defense against the risk of wash-out. Pan treeholes, in particular, are subject to rapid overflow during heavy rains, whereas sheltered rot-hole cavities are not. My student Marco Notarangelo attempted to pursue this question, but the project faltered due to unfavorable weather and more feasible alternative projects.
2. Slaughtering behavior by predatory Anopheles barberi fourth instar larvae. My PhD student Michelle Willey found this in her (unpublished) dissertation research on functional response of these larvae, which are voracious predators of first instar Oc. triseriatus. She found that the larvae were limited in the number of prey they could consume, as expected, but beyond that they continued to kill additional prey without eating them. These studies were done in a lab, and it would be interesting to see if such behavior also occurs in the field. The problem is getting enough A. barberi to do a good study. Also, given their nasty tendencies, what do they do to first instar A. barberi, and are these behaviors modified by other food sources?
3. Why aren’t Anopheles barberi present in peninsular Florida? And for that matter, why aren’t Oc. hendersoni found in peninsular Florida? I lack explanatory hypotheses for these questions; given the number of mosquito biologists working in Florida I am surprised that (to my knowledge) these questions have not been addressed.
B. The enigmatic Orthopodomyia
1. What is going on with this genus? During my decades of randomly slurping treehole water in Massachusetts beginning in 1980, I have only rarely encountered this genus. For many years I could only find O. alba (which may have been a state record) in only a couple of treeholes with very restricted openings, consistent with old descriptions of the species (e.g., Jenkins and Carpenter 1946). These could only be found sporadically. John Soghigian and I only began to see O. signifera in Massachusetts since about 2005, and only late in the season. These observations coincided with the occurrence of T. rutilus, which had never been seen in Massachusetts prior to 1997, and also only late in the summer. I suspect that, as Orthopodomyia larvae overwinter and are killed when treeholes freeze completely, cold winters may account for the absence of this genus earlier in the season; they are probably moving up from the south as the season progresses. Copeland and Craig (1990) reported that O. signifier are susceptible to freezing, whereas O. alba larvae are freeze tolerant.
2. Are O. alba and O. signifera really different species, or is alba simply a color variant? The adults are indistinguishable. I have not delved into the original species descriptions, but some DNA sequencing would be of interest (if only to a few). If truly separate, have they originated sympatrically? These species will not be easy to study as they haven’t been colonized in the lab and refuse to bite mammals as far as I know.
C. Toxorhynchites
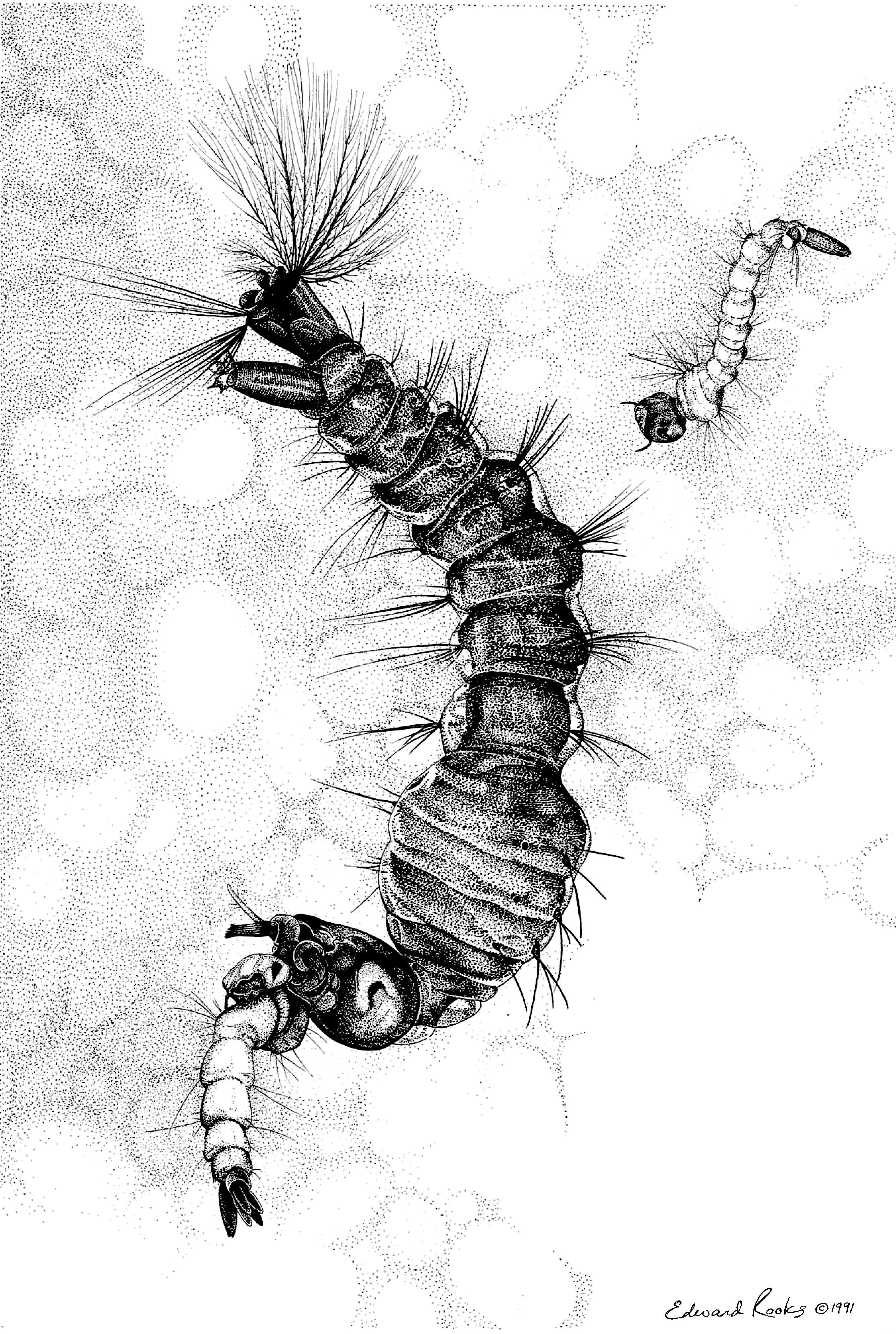
Figure 7. Toxorhynchites rutilus consuming a larva of Ochlerotatus spp. Drawing by Edward Rooks.
1. While trying to keep a daily track of egg laying by Toxorhynchites in about 10 treeholes one fall, during a period of heavy rains, eggs (which float on the water surface) were disappearing when treeholes were at capacity. Might they also show a similar preference for sheltered habitats, similar to Anopheles barberi? They are certainly not difficult to find in open habitats, but a preference for sheltered ones would not be surprising.
2. The above phenomenon may also account for the disappearance of Toxorhynchites in south peninsular Florida after a period of successive hurricanes, ca 2004. I’ve only heard of this anecdotally and I’d be interested to know if Toxorhynchites has made a comeback since those hurricanes.
3. Climate change appears to have affected Toxorhynchites distribution, as it was not seen in Massachusetts until the late 1990s when my PhD student John Dennehy discovered it (Dennehy and Livdahl 1999) and it has made consistent appearances in late summer since then. My PhD student John Soghigian was keeping some records of winter temperature minima and the timing of Toxorhynchites’ arrival in central Massachusetts, although those data may not have survived his several postdoctoral relocations. A more careful study of this species’ arrival each year in relation to weather data may be revealing.
D. Terrestrial species—slugs, ants, mites
Aedes, Ochlerotatus and Orthopodomyia eggs may need to sit on the sides of their treeholes or other containers for months at a time, during which they present a source of potential nutrition to terrestrial species which are often seen crawling around the water margin. These include ants, pulmonate snails and slugs. What are the effects of these species on egg survival? I have yet to find a study that addresses these potential interactions. Ants could be excluded from containers with tanglefoot, and mollusks could simply be manually removed. In addition to the potential effects of slug grazing, the influence of a coating of slug slime on egg hatch may be of interest.
Mite-egg interactions-- same as above, no studies I’ve heard of, although there have been some observations of mites on adult mosquitoes, possibly using them for transport. We have seen many mites on our egg papers over the years. As some mites can pierce and consume fly eggs on carrion (see, e.g., Wilson and Knollenberg 1987), mites could have important effects on mosquito populations. But really— who wants to work on mites, which all look the same?
III. The Aedes albopictus invasion of the Americas.
This species rapidly occupied most of the gulf coast region of the southeastern US, especially within domestic containers such as tires, which in that region had long been dominated by Ae. aegypti (see, e.g., Figs. 8, 9). Many investigators have presumed that this takeover has resulted from competitive superiority of Ae. albopictus, over aegypti, although the speed of takeover, as shown by our studies in Bermuda (Kaplan et al. 2010) which showed the extinction of aegypti within just a few years (Fig. 10), suggested that other types of interactions could have played a part, and results have conflicted about whether larval competition is sufficiently intense to account for such dominance (e.g., Kaplan et al. 2010, Juliano 1998).
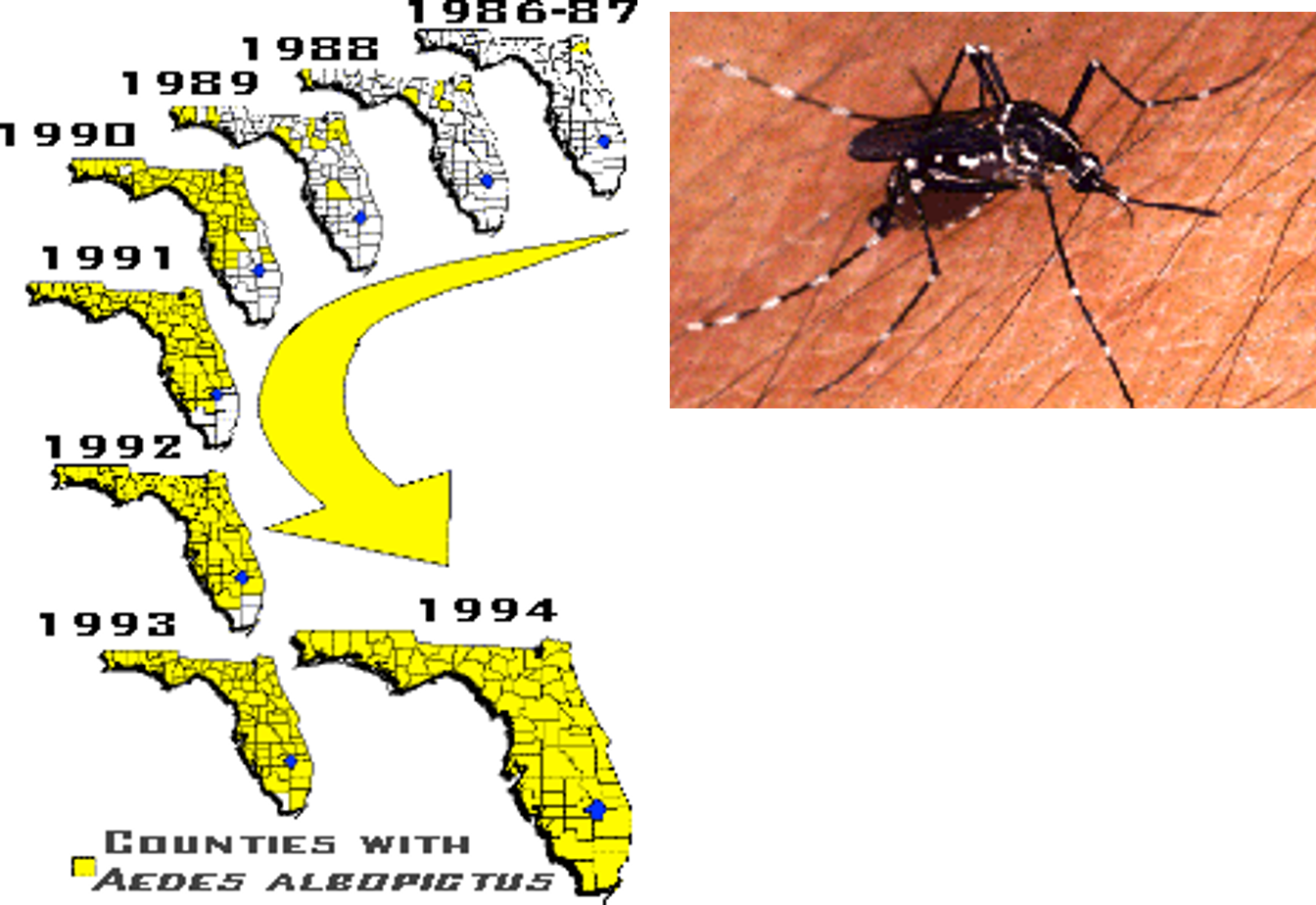
Figure 8. The advance of Aedes albopictus in Florida. Yellow counties mark the first record of this species, beginning with its discovery in Jacksonville in 1986. Thanks to George O’Meara, Florida Medical Entomology laboratory.
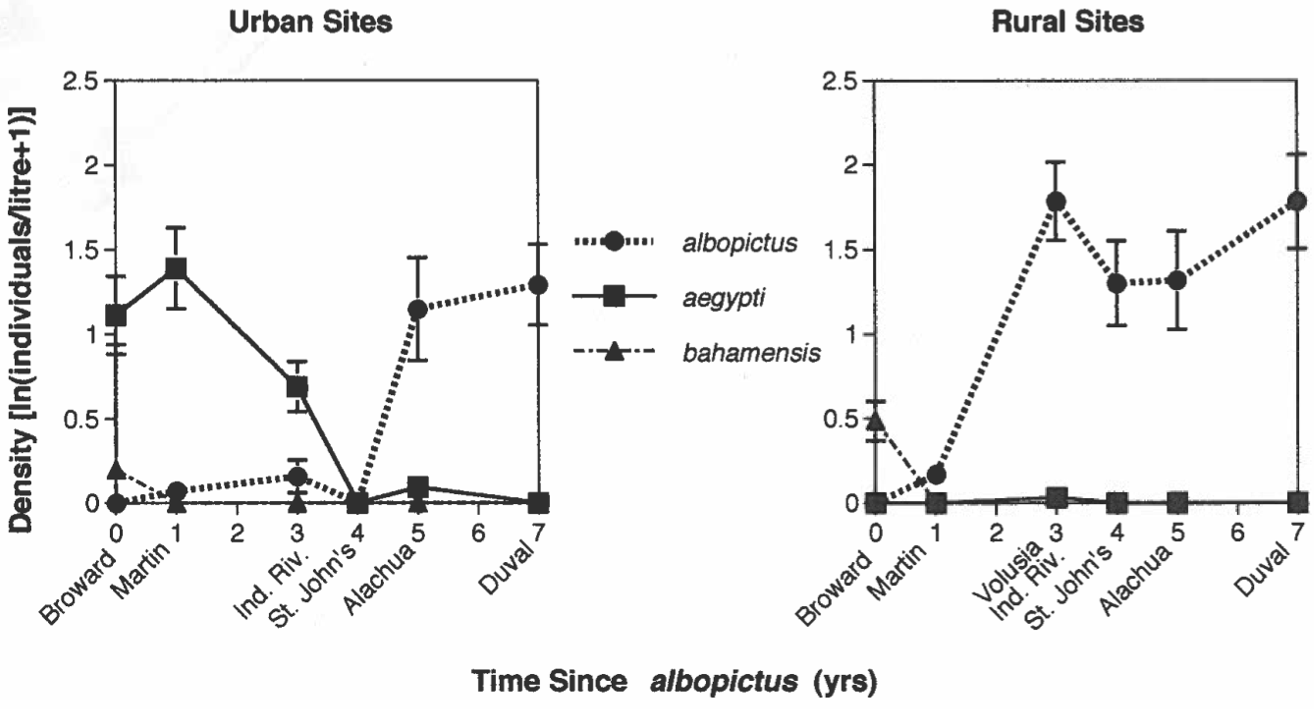
Figure 9. Data from samples taken from tires found in urban (left) and rural )(right) sites in Florida, 1993. The horizontal axis indicates the number of years that had elapsed since Ae. albopictus was first reported in each county. From Berman and Livdahl, unpublished data.
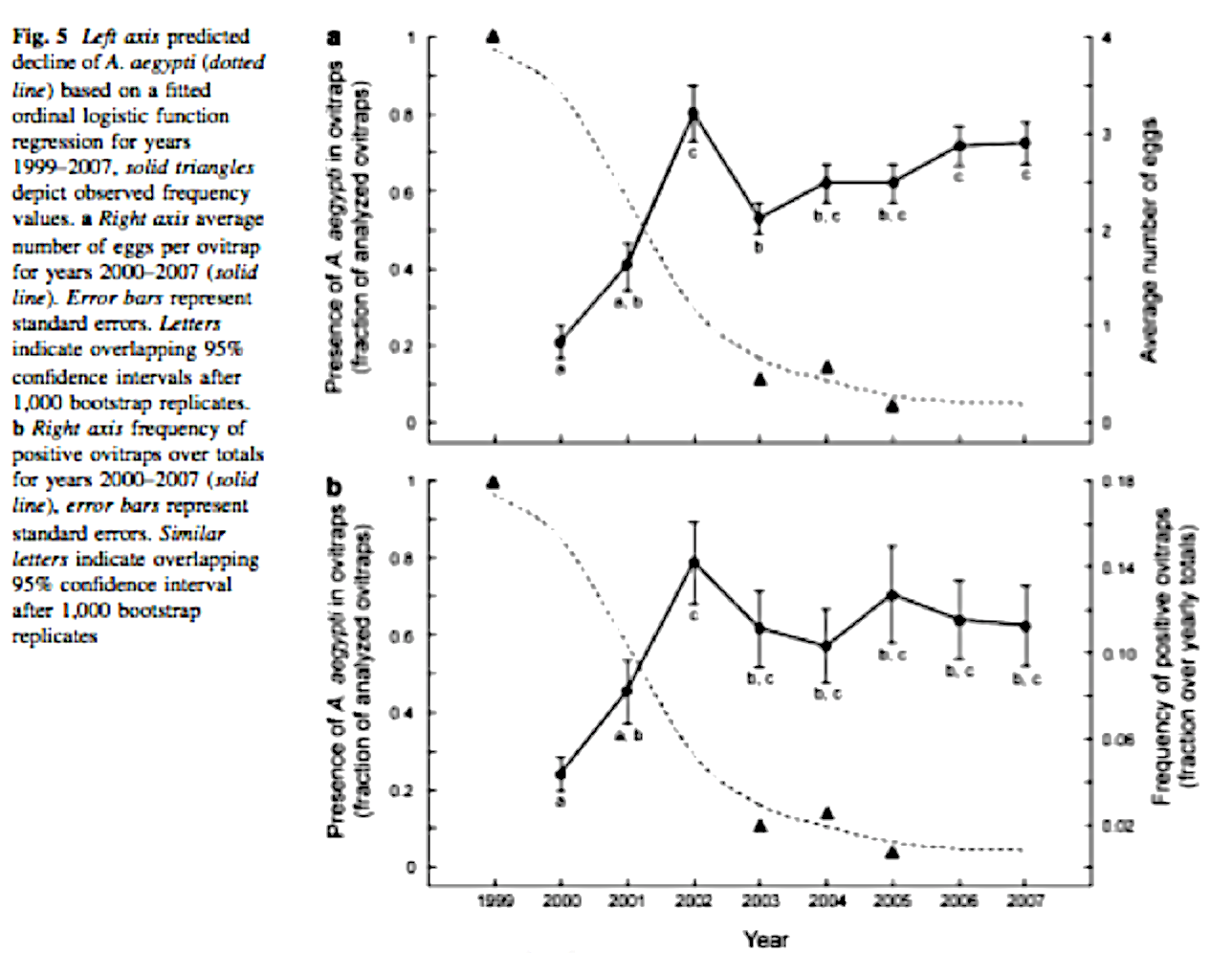
Figure 10. The takeover of Bermuda by Aedes albopictus. Egg counts are from ovitraps deployed by the Bermuda Vector Control unit, which were identified to species by Kaplan et al. (2010).
A. The possible role of egg hatch inhibition, and differential susceptibility of Ae. aegypti and Ae. albopictus eggs in this invasion. My colleagues and I found that albopictus hatching is only suppressed to a certain degree by larval density, while aegypti hatching can be suppressed entirely in laboratory containers (Fig. 11, Edgerly et al. 1993). My former BA honors student Nate Clements performed experiments which suggested that susceptibility of eggs to larval inhibition has an inherited basis in Ae. albopictus, although the study was not extensive enough to exclude maternal effects.
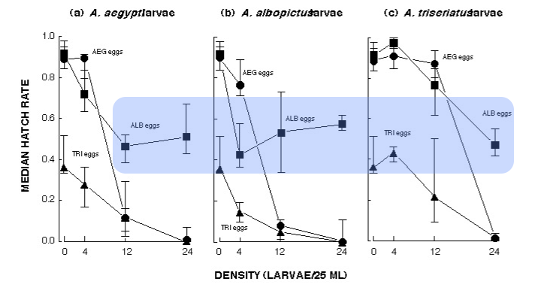
Figure 11. Egg hatch responses of three mosquito species to the density of fourth instar larvae within 30 ml vials (pooled across all three species). From Edgerly et al. (1993). Note the resistance of albopictus hatch response to high densities of larvae.
In addition to a more rigorous genetic study, this interaction needs more attention in the field, as it may help to understand the rapid displacement of aegypti. With only one exception (Ponnusamy et al. 2011), to my knowledge nobody outside of my lab has looked at this. Ponnusamy et al. (2011) showed that bacteria have direct stimulatory effects on egg hatch in addition to their indirect effects on dissolved oxygen, previously thought to be the primary hatch stimulus. Grazing of egg surface bacteria by larvae remains the presumptive effect that larvae impose on eggs (Edgerly and Marvier 1992). A challenge remains to determine whether this interaction has influenced population dynamics.
B. Feeding suppression by male Aedes albopictus harassing female Aedes aegypti. The laboratory study by my students Soghigian et al. (2014) was stimulated by a single observation (Livdahl, pers. obs.) as I sat alone in West Virginia woodlot. I was attempting to collect biting Ochlerotatus triseriatus females, and as I watched one approaching me, it was deterred from its approach by the advances of what appeared to be a male Aedes albopictus. Aedes albopictus males have the distinctive behavior of being attracted to blood sources, where they are likely to find mates as they come to feed. We found that the presence of male Ae. albopictus suppressed not only bloodfeeding, but also feeding on sugar water by Ae. aegypti (Fig. 12, 13). The converse effect of Ae. aegypti males on Ae. albopictus female feeding did not occur to a significant extent.
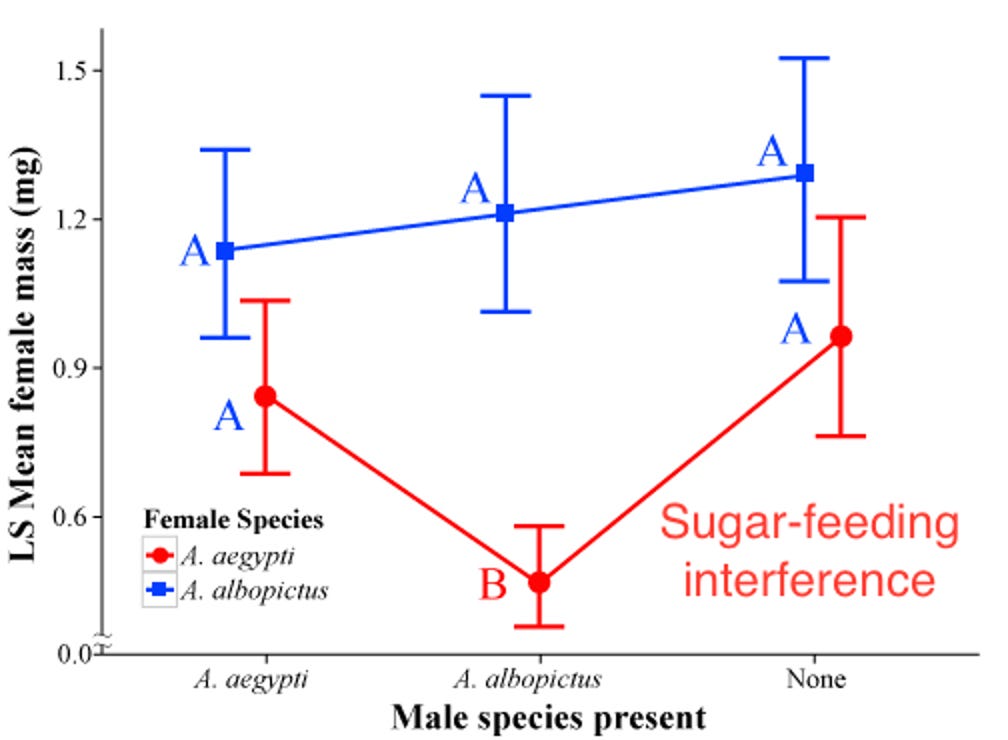
Figure 12. Sugar-feeding success of Aedes aegypti and Ae. albopictus females in laboratory cages, provided with sugar water solution and exposed to males of either species or in the absence of males. Relative feeding success is indicated by mass of females after 24 h, adjusted for female size. From Soghigian et al. (2014)
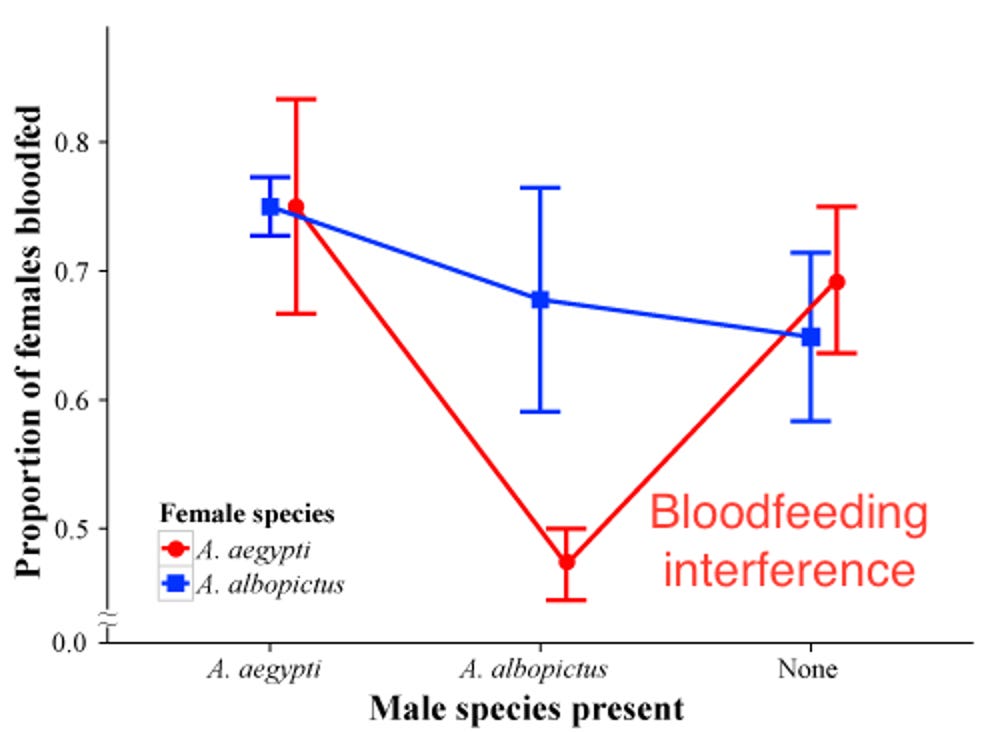
Figure 13. Blood-feeding success of Aedes aegypti and Ae. albopictus females in laboratory cages, provided with access to a laboratory mouse and exposed to males of either species or in the absence of males. Relative feeding success is indicated by mass of females after 24 h, adjusted for female size. From Soghigian et al. (2014)
Similar suppression has since been found with Ae. albopictus males affecting Ae. koreicus blood feeding (Ciochetta et al. 2023). Zhang et al. (2024) demonstrated the suppressive influence of high male:female sex ratios on feeding in conspecific trials using both Ae. albopictus and Ae. aegypti, indicating the potential importance of male harassment behavior in sterile male release programs. The inhibition of feeding by females has obvious consequences, not only to female reproduction but also to survival, and this interaction merits some serious attention in field situations, if possible. As an aside, we were impressed in our study by the ability of laboratory mice to consume many biting females.
C. Mating interactions between Aedes aegypti and Aedes albopictus. Early work of Gubler (1970) and Nasci et al. (1989) suggested that albopictus males are capable of mating interference with aegypti. These were not given much credence initially, but have been supported by the work of Phil Lounibos’ group, including their demonstration that albopictus males satyrize females of aegypti by releasing a seminal protein that sterilizes aegypti females (Tripet et al. 2011). This provides a fascinating explanation of the. rapid takeover of domestic container habitats once dominated by aegypti. It also raises interesting questions:
1. Why has albopictus not displaced aegypti in the southern hemisphere? Although the North American and Caribbean introductions have come from temperate Japan, while Brazilian South American introductions have tropical Asian origins (Birungi and Munstermann 2002, Kotsakiozi et al. 2017), it is curious that the tropical populations of albopictus did not acquire the genes necessary for similar dominance of aegypti, which has been sympatric with albopictus in tropical habitats for centuries. Several experiments have looked at competition between these species in South America, finding that larvae of the two species can compete, but no dramatic reduction in Ae. aegypti has been reported. Results from an experiment and analysis by Lizuain et al. (2022) suggest that the two species can coexist competitively, consistent with our own findings from a Florida competition experiment (Kaplan et al. 2010).
Honorio et al. (2017) exposed Ae. albopictus males from mainland Florida and also from Brazil to virgin female Aedes aegypti from Key West (which had not been exposed to Ae. albopictus) and to females from Brazil, and found that the Brazilian males were relatively unsuccessful at inseminating Ae. aegypti females. Thus, there is a behavioral component to satyrization as well as a biochemical one.
2. Given the potential for rapid evolution in these species, it may be fruitful to look for the following:
a) Evolution of resistance to the sterilizing protein by aegypti in North American habitats. This would be an intensely molecular project, well beyond my pay grade.
b) Some sort of divergence has occurred due to selection in favor of increased choosiness by aegypti females, i.e., resistance to albopictus male advances by aegypti females. Alternatively, is it reduced attractiveness of the aegypti females? Either way, such reduced cross-insemination has been demonstrated (Bargielowski et al. 2013), but to my knowledge the mechanism for this evolved choosiness/unattractiveness has not been elucidated.
Hypothesis: it’s the wingbeat frequency, stupid! Male mosquitoes have bushy antennae, while females do not. These antennae are sound receptors, and the only function males serve is to find and inseminate females. I suggest looking at each species’ responses to different sound frequencies for populations of aegypti that have been exposed to albopictus versus those that have not, and to measure the wing beat frequencies of females from such populations. I suggested this potential mechanism to a prominent mosquito ecologist, and he was highly skeptical, but this seems worth a shot for someone with access to aegypti and albopictus material from a variety of places. A crazy idea, but perhaps so crazy that it just might work. The importance of wingbeat harmonization in the mating ritual of Aedes aegypti has been shown (e.g., Cator et al. 2009), so this might not be such a wacky idea.
While considering prezygotic mechanisms that avoid hybridization, I hope someone will take a look at wing beat frequencies of female Oc. triseriatus and Oc. hendersoni in sympatric and allopatric parts of their ranges (see section I.D.), and responses of males to those frequencies. Hmm, hmm, humm.
Acknowledgments
I have many to thank for their contributions. These include: graduate students Robert Koenekoop, Michelle Willey, John Dennehy, Christopher Vitek, Camilo Khatchikian, John Soghigian, Jamie Fitzgerald and Mark Berman, Kristina Riccio, Marco Notarangelo, Matthew Chmieliewski, Matthew Warndorf, Katy Gibbs Harvey, Linda Valsdottir, Levi Seeley; honors and other undergraduate researchers David Hawkins, Michael Rosenzweig, Sarah Zapolsky, Christina Ferretti, Nathaniel Clements, Peter Morgan, Stephen Futterweit, Rachel Kaiser, Ashleigh Stanton, Heather Erb, Cynthia Sacramone, Steve Gaimari; collaborators Janice Edgerly-Rooks, Phil Robakiewicz, Deb Robertson, David Kendell, Ted Andreadis, Elizabet Estallo, Florencia Sangermano, as well as colleagues Nicholas Thompson, Phil Lounibos, George O’Meara, Steve Juliano.
References
Bargielowski IE, Lounibos LP, Carrasquilla MC. 2013. Evolution of resistance to satyrization through reproductive character displacement in populations of invasive dengue vectors. PNAS 110 :2888-2892. https://doi.org/10.1073/pnas.121959911
Birungi J, Munstermann LE. 2002. Genetic structure of Aedes albopictus (Diptera: Culicidae) populations based on mitochondrial ND5 sequences: evidence for an independent invasion into Brazil and United States. Ann. Entomol. Soc. Am. 95(1): 125-132.
Bradshaw WE, Holzapfel CM. 1983 Predator-mediated, non-equilibrium coexistence of tree-hole mosquitoes in southeastern North America. Oecologia (Berlin) 57: 239-256.
Cator LJ, Arthur BJ, Harrington LC, Hoy RR. (2009). Harmonic convergence in the love songs of the dengue vector mosquito. Science 323, 1077–1079. 10.1126/science.1166541
Ciocchetta S, Frentiu FD, Montarsi F, Capelli G, Devine GJ. (2023) Investigation on key aspects of mating biology in the mosquito Aedes koreicus. Medical and Veterinary Entomology, 37(4), 826–833. https://doi.org/10.1111/mve.12687
Connell JH. 1961. The influence of interspecific competition and other factors on the distribution of the barnacle, Chthamalus stellatus. Ecology 42: 710–723.
Copeland RS and Craig GB Jr . 1990. Habitat segregation among treehole mosquitoes (Diptera: Culicidae) in the Great Lakes region of the United States. Ann. Entomol. Soc. Am. 83: 1063– 1073.
Copeland RS and Craig GB Jr. 1992. Interspecific competition, parasitism, and predation affect development of Aedes hendersoni and A. triseriatus (Diptera: Culicidae) in artificial treeholes. Ann. Entomol. Soc. Am. 85: 154–163.
Darsie RF, Ward RA. 2004. Identification and Geographical Distribution of the Mosquitoes of North America, North of Mexico. University Press of Florida, 400 pp.
Dennehy J, Livdahl T. 1999. First record of Toxorhynchites rutilus (Diptera: Culicidae) in Massachusetts. Journal of the American Mosquito Control Association 15: 423-424.
Edgerly JS, Marvier MA (1992) To hatch or not to hatch? Egg hatch response to larval density and to larval contact in a treehole mosquito. Ecol Entomol 17: 28–32.
Edgerly JS, Willey M, Livdahl T. 1993. The community ecology of Aedes egg hatching: implications for a mosquito invasion. Ecological Entomology 18: 123-128.
Erthal J, Soghigian J, Livdahl T. 2012. Life cycle completion of parasite Ascogregarina taiwanensis (Apicomplexa: Lecudinidae) in non-native host Ochlerotatus japonicus (Diptera: Culicidae). J. Med. Entomol. 49:1109-1117. DOI: 10.1603/ME12018
Fitzgerald, J. and T. Livdahl. 2019. Vertical habitat stratification in sympatric and allopatric populations of Aedes hendersoni and Aedes triseriatus (Diptera: Culicidae). Journal of Medical Entomology 56:311-319. DOI: 10.1093/jme/tjy107.
Fitzgerald J, Livdahl T. 2023. Vertical habitat stratification in Aedes triseriatus and Aedes hendersoni (Diptera: Culicidae): complications associated with sloped and flood-prone landscapes. Journal of Vector Ecology 48(2), 113-123. https://doi.org/10.52707/1081-1710- 48.2.113
Gubler DJ. 1970. Competitive displacement of Aedes (Stegomyia) Polynesiensis Marks by Aedes (Stegomyia) albopictus Skuse in laboratory populations. J. Med. Entomol. 7::229-35. doi: 10.1093/jmedent/7.2.229.
Honorio N, Carasquilla MC, Bargielowski IE, Nishimura N, Lounibos LP. 2017. Male origin determines satyrization potential of Aedes aegypti by invasive Aedes albopictus. Biological Invasions. 20. 10.1007/s10530-017-1565-3.
Jenkins, DW, SJ Carpenter. 1946. Ecology of the tree hole breeding mosquitoes of nearctic North America. Ecol. Monogr. 16:31-47.
Juliano S. 1998. Species introduction and replacement among mosquitoes: interspecific resource competition or apparent competition? Ecology 79:255-268. 10.2307/176880.
Kaplan L, Kendell D, Robertson D, Livdahl T, Khatchikian C. 2010. Aedes aegypti and Aedes albopictus in Bermuda: extinction, invasion, invasion and extinction. Biological Invasions 12:3277-3288.
Kotsakiozi P, Richardson JB, Pichler V, Favia G, Martins AJ, Urbanelli S, Armbruster PA, Caccone A. 2017. Population genomics of the Asian tiger mosquito, Aedes albopictus: insights into the recent worldwide invasion. Ecol. Evol. 24:10143-10157. doi: 10.1002/ece3.3514.
Lizuain AA, Maffey L, Garzón M, Leporace M, Soto D, Diaz P, Salomón OD, Santini MS, Schweigmann N. 2022. Larval Competition Between Aedes albopictus and Aedes
aegypti (Diptera: Culicidae) in Argentina: Coexistence and Implications in the Distribution of the Asian Tiger Mosquito, Journal of Medical Entomology 59:1636–1645. https://doi.org/10.1093/jme/tjac102
Lounibos LP, Machado-Allison CE. 1983. Oviposition and egg brooding by the mosquito Trichoprosopon digitatum in cacao husks. Ecol. Entomol. 8:475-478.
Nasci RS Hare SG, Willis FS. (1989) Interspecific mating between Louisiana strains of Aedes albopictus and Aedes aegypti in the field and in laboratory. J. Amer. Mosq. Contr. Assn. 5: 416– 421.
Pianka, E. 2010. Evolutionary ecology, 6th edition. Benjamin Cummings, San Francisco. 512 pp.
Ponnusamy L, Böröczky K, Wesson DM, Schal C, Apperson CS (2011) Bacteria Stimulate Hatching of Yellow Fever Mosquito Eggs. PLOS ONE 6(9): e24409.https://doi.org/10.1371/journal.pone.0024409
Ray R, Nakata T, Henningsson P et al. 2016. Enhanced flight performance by genetic manipulation of wing shape in Drosophila. Nat Commun 7, 10851 (2016). https://doi.org/10.1038/ncomms10851.
Soghigian J, Gibbs K, Stanton A, Kaiser R, Livdahl T. 2014. Sexual harassment and feeding inhibition between two invasive dengue vectors. Envir. Health Insights 2014:Suppl. 2 61-66. doi: 10.4137/EHI.S16007.
Soghigian J, Livdahl T. 2021. Field evidence of mosquito population regulation by a gregarine parasite. Journal of Medical Entomology 58:1188–1196, https://doi.org/10.1093/jme/tjab009
Tripet F et al. 2011. Competitive reduction by satyrization? Evidence for interspecific mating in nature and asymmetric reproductive competition between invasive mosquito vectors. Am J Trop Med Hyg 85, 265–270.
Wilson DS, Knollenberg WG. 1987. Adaptive indirect effects: the fitness of burying beetles with and without their phoretic mites. Evol Ecol 1, 139–159. https://doi.org/10.1007/BF02067397
Zhang, D., Maiga, H., Li, Y. et al. Mating harassment may boost the effectiveness of the sterile insect technique for Aedes mosquitoes. Nat Commun 15, 1980 (2024). https://doi.org/10.1038/s41467-024-46268-x.